





Mononuclear Phagocytes Produce Key Inflammatory Cytokines and Chemokines
- Mononuclear phagocytes (monocytes and macrophages) are essential for the development of inflammation and together with neutrophils are the phagocytic cells involved in the clearance of inert particles and microbial agents.
- Monocytes are bone marrow-derived cells and are continuously released into the blood.
- When these cells are recruited by chemotactic molecules and leave the circulation, they become activated and differentiate into macrophages.
- Other names have been applied to tissue macrophages such as Kupffer cells in the liver, histiocytes in connective tissues, mesangial cells in the kidneys, osteoclasts and chondroclasts in bones and cartilage, alveolar macrophages in the lungs, and microglial cells in the brain.
- In contrast to neutrophils, which are end-stage nonreplicating cells continually replaced from the bone marrow, mononuclear phagocytes can proliferate in situ and are long-lived (i.e., months to years).
- Macrophages actually have a number of important functions in body defense such as
- (1) capture by phagocytosis and intracellular killing of microorganisms, infected cells, and tumor cells through PAMP recognition;
- (2) scavenging of worn-out cells, apoptotic bodies, and other debris potentially harmful to tissues;
- (3) processing and presentation of antigens for recognition by T cells, expressing co-stimulatory molecules, mainly during secondary adaptive immune responses;
- (4) releasing cytokines and chemokines that play a major role in innate immune responses.
- (5) regulate immunity through the alternatively activated pathway
- The major cytokines produced by macrophages are TNF-alpha, IL-1 beta, IL-6, and IL-8 and IL-33
- Some important systemic effects of cytokines produced by macrophages are fever induced by IL-1, wasting (i.e., cachexia) caused by TNF-alpha, production of acute phase proteins by liver induced by IL-6, and an increase in the maturation and release of neutrophils from bone marrow by IL-3.
- Although phagocytosis and cytokine/chemokine production are the two key effector functions of macrophages in the innate immune response, these cells also have a role in adaptive responses as antigen-presenting cells and as targets of the effector components of the cellular and humoral adaptive responses, being activated by T cell-derived cytokines and antibodies.
- Macrophage capacity to kill pathogenic microorganisms also can be overcome by microbes that are able to survive inside cells, establishing an intracellular infection, such as Salmonella spp, Mycobacterium tuberculosis, Cryptococcus neoformans, or Toxoplasma gondii.
Dendritic Cells Produce Inflammatory Cytokines and Initiate Adaptive Immunity by Presenting Antigens to T Cells
- Dendritic cells (DCs) are a sparsely distributed, migratory group of cells that are specialized to sample the sites of entry of infectious agents.
- Dendritic cells are not only important elements of the innate immune system but also constitute the most efficient antigen-presenting cells for T cell activation, thus linking innate and adaptive immune responses.
- There are different subsets of DCs, most commonly found in the systemic circulation are:
- (1) plasmacytoid (pDCs), derived from lymphoid precursors
- (2) myeloid (mDCs), derived from myeloid precursors
- A distinguishing feature of pDCs is their capacity to produce large amounts of type I interferons, i.e., IFN-γ and IFN-γ, during the course of viral infection driven by activation via the endosomal TLR7 and TLR9 pathways
- mDCs respond best to bacterial infection, recognizing LPS via the cell-surface TLR4 together with the production of IL-12 and TNF-α.
- Both types of DCs circulate in the blood prior to migration into epithelial sites of skin and mucosal tissues, where they remain as immature DCs.
- These cells utilize the mechanisms of pinocytosis (i.e., ‘‘cell-drinking,’’ a form of endocytosis in which small droplets of liquids are brought into the cell suspended within small vesicles) and phagocytosis for the uptake of a wide variety of foreign proteins and infectious agents.
- This results in the activation of DCs during which the maturation of the cells is promoted with the expression of the chemokine receptor CCR7 allowing the cells to migrate from the periphery to secondary lymphoid organs.
- During emigration, the captured molecules are processed into small peptides, linked to the grooves of MHC-II molecules, and expressed on the cell surface of DCs. By the time DCs reach secondary lymphoid organs, they are able to present antigens to populations of naive and memory T cells.
- Activated DCs also produce cytokines of the IL-12 family (IL-12, IL-23, and IL-27) with Th1-cell polarizing capacity and IL-6, IL-10, and TGF-β with regulatory T cell polarizing effects.
Mast Cell Mediators Regulate a Variety of Physiological Systems
- Mast cells (MCs) originate in bone marrow and circulate as CD34+ progenitor cells, differentiating into mature mast cells under the influence of cytokines only after entry into tissues.
- They are distributed throughout the body as resident cells, particularly in association with blood vessels and nerves, and in close proximity to mucosal surfaces that interface with the external environment
- After their development from bone marrow-derived progenitor cells that are primed with stem cell factor (SCF), mast cells continue their maturation and differentiation in peripheral tissue, developing into two well-described subsets of mast cells, MC(T) and MC(TC) on the basis on their enzyme content.
- The first subset consists of MCs containing tryptase (T), i.e., MC(T); the second are mast cells containing both tryptase (T) and chymase (C), i.e., MC(TC).
- The TC type predominates in normal skin and intestinal submucosa, and contains tryptase, chymase, a cathepsin G-like protease, and a carboxypeptidase, whereas the tryptase-containing T type MC(T) is found in intestinal mucosa and lung alveolar wall.
- Mast cells are particularly important in innate immune responses because they are capable of detecting infectious agents and initiating an acute inflammatory response through the secretion of mediators.
- These cells recognize microbial PAMPs through TLR1, -2, -4, and -6 and the complement-derived molecules iC3b, and anaphylatoxins (C4a, C3a, and C5a) through their respective receptors.
- After activation, mast cells immediately extrude granule-stored preformed mediators (histamine, tryptase, chymase, and carboxypeptidase) and, in minutes, generate lipid-derived prostaglandin D2 (PGD2), and leukotrienes LTC4, LTD4, and LTE4)
- Late, within hours, they synthesize chemokines (CCL3, CCL4, and CCL5) and cytokines (TNF-α, IL-4, IL-5, and IL-6).
- The rapid release of these mediators promotes vascular permeability, induces vasoconstriction, and recruits eosinophils, neutrophils, and other cells within a very short time.
Basophils and Eosinophils Also Participate in Innate Immunity
- Basophils are granulocytes derived from bone marrow precursor cells and released into the blood circulation.
- Their characteristics and functions are similar to those described for mast cells.
- Eosinophils are another kind of blood granulocyte that can be recruited to sites of innate immune reactions, where their number can be 100 times higher than in the blood.
- Eosinophil cytoplasmic granules contain a variety of cationic proteins that exert several biological effects on normal cells and infectious agents, and these include: major basic protein (MBP), eosinophil cationic protein (ECP), eosinophil-derived neurotoxin (EDN), and eosinophil peroxidase (EPO).
- During the innate immune responses, and induced by phagocytosis of opsonized particles, the eosinophil granule content is released, acting mainly on extracellular helminthic parasites and contributing to tissue damage in inflammatory diseases.
- Other eosinophil products also participate in acute and chronic inflammatory reactions, particularly in allergic diseases (see Chapter 18, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012).
- These include LT, PAF, and cytokines such as granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-3, IL-5, IL-6, IL-8, TNF-α, and TGF-β.
Natural Killer Cells Recognize and Kill Virus-Infected Cells
- Natural killer (NK) cells are large granular lymphocytes derived from bone marrow precursors and are found mainly in peripheral circulation (5 to 20 percent of total lymphocytes), spleen, liver, and bone marrow.
- Although NK cells belong to the lymphoid lineage, they express neither T (TCR/CD3 complex) nor B (CD19) cell surface markers and are considered the third most important lymphocyte subset (see Chapter 2, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012).
- NK cells have a morphology similar to that of activated cytotoxic lymphocytes, i.e., a large size, an abundant endoplasmic reticulum (ER), and the presence of preformed granules containing perforins and granzymes.
- Because they express CD16 (Fc-yRIII) and CD56, NK cells are usually identified as CD3-/CD56+/CD16+ cells.
- The role of NK cells in the innate immune response is remarkable because of their nonspecific capacity to eliminate target cells, e.g., virus-infected or malignant cells, through apoptosis independently of T or B cells.
- NK cells also stimulate inflammatory responses through secretion of IFN-gamma, TNF-alpha, GM-CSF, and chemokines (CCL4, CCL5, CCL22) (see Chapter 9, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012).
- NK cells recognize and kill target cells by apoptosis basically in two ways using an array of different cell surface receptors.
- (1) They utilize the complex balance between activating and inhibitory signals that the NK cell generates to either activate or inhibit their killing activity.
- Some NK cells have receptors generically called killer inhibitory receptors (KIRs) that belong to at least two groups of unrelated molecule structures (C-type lectin and immunoglobulin superfamily), which recognize MHC-I molecules on normal self cells and which, in the normal state, inhibit activation and killing due to the presence in their intracytoplasmatic tail of immunoreceptor tyrosine-based inhibitory motifs (ITIMs) (see Chapter 5, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012).
- Another diverse set of lectin-type receptors, generically referred to as killer activating receptors (KARs), detect other molecules on altered target cells, e.g., malignant or virus-infected cells, which can provide an activation signal for the NK cell via immunoreceptor tyrosine-based activation motifs (ITAMs).
- If a target cell has decreased expression of MHC-I (see Chapter 20, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012), the inhibitory signal from KIR will be decreased, shifting the signal balance towards activation of the NK cell and initiating the process of target cell death by apoptosis.
- Once the signal balance is tipped toward activation rather than inhibition, the killing mechanism is initiated, and the NK cells utilize their intracellular granules perforin and granzymes to kill the target cells by inducing apoptosis, shown in Figure 1 (see video below).
- Upon release, perforin in close proximity to a cell slated for killing, first forms pores in the cell membrane of the target cell through which the granzymes and associated molecules then enter, inducing apoptosis (see Chapter 9, Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012).
- 2) The second method whereby NK cells kill target cells is through a hybrid mechanism in which the IgG molecule links the innate and adaptive immune systems and is referred to as antibody-dependent cellular cytotoxicity (ADCC, depected in Figure 1)
- In this scenario, the NK cell recognizes target cells to which IgG antibody has been attached through its Fab regions.
- At the same time, the Fc portion of the IgG antibody engages the NK cell through its Fc receptor referred to as CD16 (Fc-gRIII), thus linking the NK cell with the target cell.
- Following this linkage, the NK cell can destroy the target cell through induction of the apoptotic pathway.
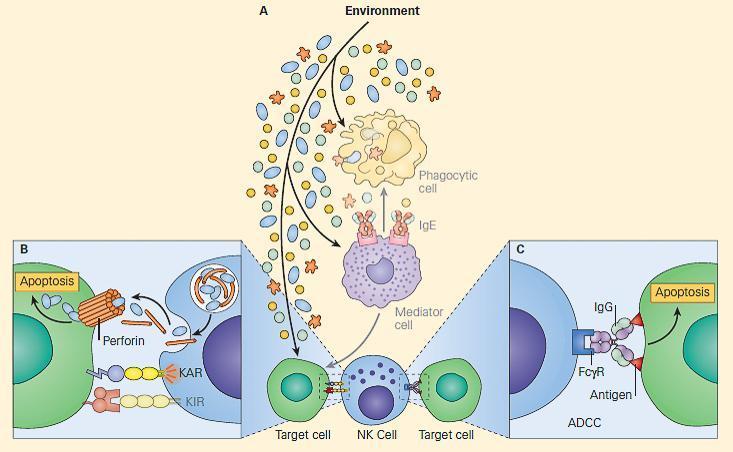
Figure 1. Panel A: Schematic representation of the NK cell and its role in innate and adaptive immunity showing the two ways it can kill target cells. Panel B: Shows direct killing when KAR > KIR, during which release of perforins and granzymes lead to cell death by apo-ptosis (innate immunity). Panel C: Shows killing by ADCC by utilizing an immunoglobulin molecule to bridge the NK cell with a target cell (adaptive immunity). KAR = killer activating receptor; KIR = killer inhibitory receptor. [Reproduced with permission from Bellanti, JA (Ed). Immunology IV: Clinical Applications in Health and Disease. I Care Press, Bethesda, MD, 2012].
Natural killer T (NKT) cells
- Another subset of natural killer cells has been identified as NKT cells, which were originally defined by the co-expression of T cell markers (CD3/TCR complex) along with characteristic surface receptors for NK cells (CD56, CD58, and CD161), indicating a dual nature of this subset.
- NKT cells can either express or not express CD4 or CD8 molecules and, similar to NK cells, are considered part of the innate immune response because they act swiftly during infections, killing microorganisms or cells and producing cytokines without the need of slower differentiation or proliferative processes as with T and B cells.
- The original definition of NKT cells, which included a broader group of cells, is now restricted to a more specific subset, the so-called invariant NKT (iNKT) cells.
- They characteristically express on their surface a TCR invariant or constant region (Vα24/J18/Vβ11 in the human) able to recognize glycolipids presented on CD1d molecules
- The CD1 family (CD1a through CD1d) consists of antigen-presenting molecules encoded by genes located outside of the MHC and structurally similar to the MHC-I molecules.
- In comparison to classical MHC molecules, the CD1 antigen-binding groove is highly hydrophobic and adapted for the presentation of lipid antigens that consist largely of glycolipids.
- The cognate α-galactosylceramide (α-Gal Cer, derived from a marine sponge) is the model glycolipid recognized by iNKT cells.
- iNKT cells are present in internal organs, including the thymus, bone marrow, liver, and spleen.
- There are other populations of cells expressing T and NK membrane markers differing from the iTCR cells, but showing variable TCRs, which are able to recognize a broader set of structures in microorganisms, and these are generically called NKT-like cells.
Mucosa-associated invariant T (MAIT) cells
- These cells are found in low numbers in the intestinal lamina propria of humans and require commensal flora to expand and/or persist.
- They express a semi-invariant TCR (Vα7.2-Jp33 chain), are double negative CD4- and CD8-, recognize hydrophilic antigens presented in MR1 (MHC-I-related) molecules present in B cells, and upon antigen activation, produce IFN-, IL-4, IL-5, and IL-10.
- MAIT cells have been shown to be implicated in immune responses to TB in the lung
B-1 cells
- B-1 cells are a phenotypically and functionally distinct subset of B cells, which spontaneously secrete ‘‘natural’’ IgM antibodies in the absence of any apparent stimulation by specific antigens (see Chapter 2).
- Accordingly with the expression of CD5 molecules, B-1 cells are subdivided into B-1a that carry the CD5 molecule and B-1b that do not
- B-1a cells represent the majority of B-lineage cells during neonatal life but decline thereafter.
- The repertoire of natural antibodies is much more restricted than those produced by conventional B cells and a large proportion are poly-reactive to phylogenetically conserved structures such as nucleic acids, heat shock proteins, carbohydrates, and phospholipids.
- The antibodies produced by B-1 cells may participate as a bridge between innate and adaptive immunity and make an optimal transition between the two immune responses by producing the first wave of antibodies required for antigenic clearance of viruses, bacteria, and certain parasites.
- Other functions performed by these cells are in the immune regulation through the synthesis of IL-10 and in the clearance of senescent and apoptotic cells.
- It has been suggested that they are also involved in autoimmunity as increases in CD5+ B cell frequency have been reported in patients suffering from rheumatoid arthritis, Sjögren’s syndrome, myasthenia gravis, insulin-dependent diabetes mellitus, and Hashimoto’s thyroiditis (see Chapter 19).
Marginal zone B (MZ B) cells
- MZ B cells are a rare non-recirculating subset of mature peripheral B cells exclusively located in the spleen, different from the more abundant recirculating follicular B cell subset.
- The MZ B cell population is separated from the B cell follicle by the marginal sinus.
- MZ B cells express high levels of CD1d and CD21 molecules.
- They are quick and easily activated by low levels of antigen, are potent antigen-presenting cells for naive T cells, and produce short-lived IgM antibody-forming cells involved in the early defense against blood-borne pathogens, as well as in autoreactive B cell responses.



