





The Rv144 trial demonstrated that IgG targeting of specific HIV Envelope protein (Env) regions, in particular the V2 (Haynes et al. , Gottardo et al. 2013), but also the V3 region (Gottardo et al. 2013) was correlated with protection from HIV acquisition. Therefore, understanding the factors that influence the vaccine-induced IgG recognition of certain HIV Env regions might aid in an improved and targeted HIV vaccine design.
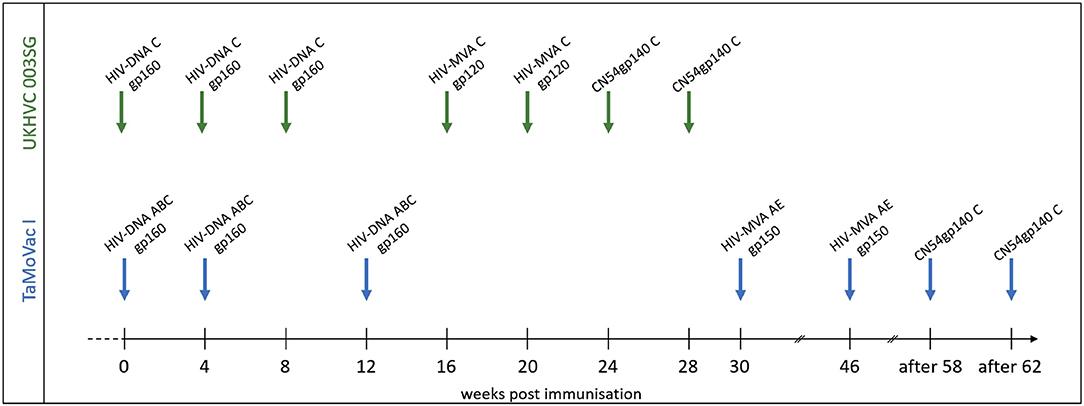
To this end, Nadai et al., closely mapped the IgG response towards Env, elicited by two different HIV vaccination strategies applied in the preventive HIV vaccine trials UKHVC003 (UK) and TaMoVac01 (Tanzania). Both trials used three DNA immunisations, followed by boosting with two recombinant Modified Vaccinia Virus Ankara (MVA), and finally two boosts with glucopyranosyl lipid adjuvant (GLA) adjuvanted trimeric CN45gp140 protein. The main differences between these trials lay in the immunogen design of the MVA, where cells infected by UKHVC003 MVA secreted monomeric gp120 protein and TaMoVac01 MVA infected cells expressing membrane bound functional gp150 protein. Further, the UKHVC003 vaccines were based on clade C only, while TaMoVac01 vaccines were based on clades A, C, B, and CRF01AE.
Fine mapping of the Env IgG recognition was carried out using a peptide microarray consisting of 15mer peptides covering the HIV-1 Env, designed to represent recently transmitted global HIV strains and the immunogen sequences used in both trials. In addition, hot spots of IgG recognition were covered by additional peptide variants. Apart from a strong response against the V3 region, the two vaccine trials differed strongly in the elicited IgG response against Env regions. Post MVA UKHVC003 IgG responses mostly targeted regions within the inter-gp-120 interface while TaMoVac01 IgG responses were dominated by a V2 and gp41 response. After the final boost, the regions targeted by the IgG responses in the TaMoVac01 vaccinees were more comparable to the UKHVV003 vaccinees, however, the V2 and gp41 response still dominated. Cross-recognition of antigenic variants of the V3 tip was not improved by the diverse immunogen sequences in TaMoVac01 over the clade C-only immunogen sequences in UKHVC003. This study shows, that structural features, but also variations in the sequence of the immunogens, might contribute to varying IgG responses elicited by varying vaccines.
Journal Article: Nadai et al. 2019. Envelope-Specific Recognition Patterns of HIV Vaccine-Induced IgG Antibodies Are Linked to Immunogen Structure and Sequence. Front. Immunol.
Article by Kathrin Held



